

I łaciate i mroziaste…
Ponad 150 lat temu pewien Austriak, augustianin z Brna na Morawach – Grzegorz Mendel, odkrył podstawowe prawa genetyki robiąc doświadczenia na grochu (Pisum).
Gregor Johann Mendel o swoich badaniach informował w latach 1865–66. Jednak zostały one zapomniane prawie na 50 lat, gdyż większe zainteresowanie budziły wówczas jego prace z zakresu meteorologii. W 1900 roku szerzej nieznane wówczas wyniki tego zakonnika potwierdziło niezależnie trzech uczonych: Hugo de Vries, Carl Correns i Erich Tschermak, co zapoczątkowało powstanie nowej nauki – genetyki. Mendel pisał o „czynnikach dziedziczenia”, bo nieznane były wówczas takie terminy jak gen czy allel.
Prawa Mendla
Istnieją dwa prawa – pierwsze i drugie, choć w obecnej formie nie zostały tak sformułowane przez ich autora.
Pierwsze prawo Mendla dotyczy czystości gamet, czyli komórek, które na każdą cechę posiadają jeden allel z pary alleli danego genu. Allel, to jedna z wersji tego samego genu, zajmujący taką samą pozycję (locus) w danym chromosomie. Każda komórka płciowa posiada tylko jeden allel z każdej pary alleli.
Drugie prawo Mendla odnosi się do niezależnej segregacji, co oznacza, że różne cechy ulegają segregacji niezależnie od siebie. Przypadek powoduje, który allel z pary genów determinujących jedną cechę będzie w gamecie w parze z innym allelem warunkującym drugą cechę, co odzwierciedli się w fenotypie, np. w kolorze oczu, włosów, sierści itp. Oznacza to, że geny jednej pary alleli są dziedziczone niezależnie od genów drugiej pary. Wobec tego w pokoleniu F2 (drugim pokoleniu potomnym) występuje rozszczepienie fenotypów w stosunku 9:3:3:1. Załóżmy, że jakaś cecha jest determinowana dwoma genami, jednym dominującym – oznaczonym wielką literą, np. A, powodującym pojawienie się np. koloru brązowego i drugim recesywnym, oznaczonym małą literą a, determinującym kolor niebieski. Geny pochodzą zawsze w połowie od matki i od ojca. Jeżeli spotkają się dwie gamety – męska i żeńska – AA i AA (osobniki brązowe), powstanie osobnik o genotypie AA, czyli homozygota dominująca brązowa. W przypadku konfiguracji AA (brązowy) i aa (niebieski) nowy osobnik będzie też brązowy, ale będzie heterozygotą dominującą (Aa lub aA) – dominacja zupełna. Jeżeli skojarzymy osobniki Aa (lub aA) i aa, można będzie uzyskać następujące potomstwo: Aa (lub aA) i aa, czyli albo brązową heterozygotę dominującą (Aa lub aA), albo homozygotę recesywną niebieską (aa). Jeżeli skojarzymy osobniki o genotypie aa i aa, całe potomstwo będzie niebieskimi homozygotami recesywnymi (aa).
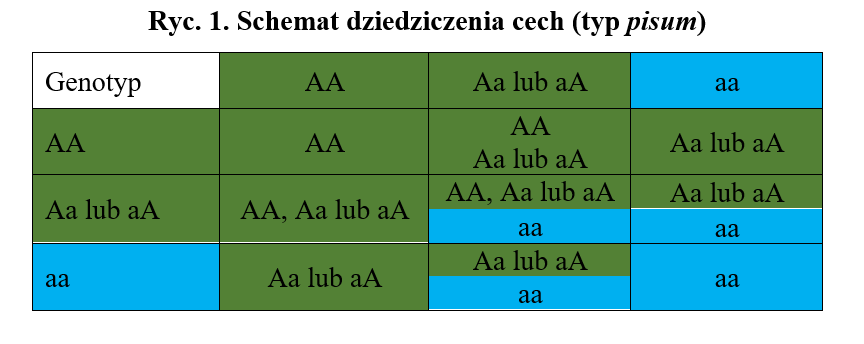
Oczywiście, nie zawsze wszystko jest takie proste.
Co odkrył Thomas Morgan?
Thomas Morgan częściowo obalił teorię Mendla, gdyż udowodnił, że geny położone w tym samym chromosomie dziedziczą się razem. Zazwyczaj jedna cecha jest warunkowana przez wiele alleli, a nie tylko przez jeden. Poza tym, allele mogą być kodominujące, np. tak jest w przypadku grup krwi u ludzi – A i B, które po połączeniu dają grupę AB, posiadającą oba antygeny widoczne na powierzchni komórek. Istnieje jeszcze dominacja niezupełna, która dotyczy heterozygot, u których ujawniły się cechy pośrednie pomiędzy allelami. Ponadto w puli genów populacji zwykle istnieje wiele alleli tego samego genu o zróżnicowanym porządku recesji, dominacji i kodominacji. Kodowanie białek uszkodzonych lub ich niekodowanie jest uwarunkowane istnieniem genów recesywnych.
Łaciate, jednokolorowe, białogrzbiete…
Umaszczenie bydła jest cechą rasową i przy istnieniu ponad 400 ras jest dość zróżnicowane, na co wpływ miało udomowienie tych zwierząt ponad 8–9 tys. lat temu w Azji. Jego dziedziczenie jest w typie pisum, opisanym przez Mendla, ale u jednej rasy – shorthorn, jest w typie zea (kukurydzy) z niekompletną dominacją.
Umaszczenie może być podstawowe, rozjaśnione lub łaciate. Można je też podzielić na jednolite lub niejednolite. Jednolitość jest cechą dominującą, a łaciatość – recesywną. Umaszczenie jednolite, to: białe, czarne, czerwone, żółte, siwe, sarnie, śmietankowe i brunatne. Natomiast niejednolite, to: pstre lub łaciate – czarno-białe, czerwono-białe, żółto-białe, dereszowate i białogrzbiete (dawniej boczaste). Jednak umaszczeniem podstawowym jest czarne i czerwone. Oba kolory koduje jeden gen – Extension, odpowiadający za wytwarzanie dwóch barwników – czarnej eumelaniny i czerwonej feomelaniny. Co ciekawe, w tych samych loci (liczba mnoga od locus) te dwa kolory są warunkowane aż przez trzy allele.
Umaszczenie czarne
Jest umaszczeniem podstawowym, determinowanym przez allel ED dominujący nad dwoma pozostałymi. Istnieją następujące kombinacje: EDED, EDe i EDE+ – zwierzęta są czarne. Należą tu takie rasy jak chińskie czarne bydło, angus i chiangus. Poza tym, tak warunkowany kolor występuje jako jeden z wielu u bydła holsztyńsko-fryzyjskiego oraz u takich ras jak brangus, balancer, limousin, lim-flex, highland, simental, simangus czy salers.
Umaszczenie czerwone
Kolejna maść determinowana jest przez recesywny allel e, który może występować w następujących kombinacjach: ee i E+e. Powoduje brak wytwarzania czarnego barwnika, co w efekcie daje fenotyp czerwony, jaki występuje u takich ras, jak polska czerwona, hereford, lincoln red, red angus, red poll, highland, santa gertrudis.
Umaszczenie dzikie
To trzecia barwa podstawowa determinowana przez allel E+. Jego działanie jest neutralne w połączeniu z pozostałymi dwoma allelami – ED i e. Warunkuje wówczas kolor determinowany przez allel będący z nim w parze. Jego obecność nie sprawia, że bydło będzie miało umaszczenie tura, ale u homozygoty E+E+ głowa, szyja i kończyny, czyli części oddalone od tułowia, będą miały cieniowanie takie, jak u dzikiego przodka. U tych zwierząt produkowane są oba barwniki – czarny i czerwony. Prawdopodobnie w takie oddziaływanie tego genu „zamieszany” jest inny gen, o nazwie agouti (od nazwy zwierzęcia, pol. aguti). Takie barwy można zobaczyć u europejskiego bydła ras prymitywnych oraz u bydła pochodzącego z Bliskiego Wschodu, a także u ras, takich jak szwajcarskie brązowe, stepowe, jersey i heck.
Anomalie
Bydło rasy holsztyńsko-fryzyjskiej posiada m.in. genotyp ee, co powinno determinować fenotyp czerwony. Jednak stwierdzono występowanie barwy rudej, prawdopodobnie spowodowane obecnością innego genu. Efekt ten nazwano czerwonym dominującym lub wariantem czerwonym. Ponadto u bydła tej rasy występuje zjawisko zmiany koloru – najczęściej z czerwonego na czarny, rzadziej z czarnego na czerwony, za co prawdopodobnie odpowiada gen recesywny.
Dziedziczenie typu zea
Według tego typu dziedziczone jest umaszczenie białe i czerwone u rasy shorthorn. Krzyżując osobnika czerwonego o genotypie C1C1 z osobnikiem białym o genotypie C2C2 otrzymuje się potomstwo F1 mroziaste – C1C2, mające wymieszane włosy czerwone i białe. W przypadku kojarzenia osobników mroziastych (C1C2 x C1C2) uzyska się w ¼ potomstwo czerwone (C1C1), w 2/4 mroziaste (C1C2, C2C1) i w ¼ białe (C2C2).
Łaciate holsztyńsko-fryzyjskie
Dominującą rasą bydła mlecznego w Polsce jest polska holsztyńsko-fryzyjska odmiany czarno-białej. Mniej liczebna jest odmiana czerwono-biała, choć swoimi cechami użytkowymi i produkcyjnymi nie odbiega od konkurentki. Obecnie istnieje jedna księga hodowlana bydła tej rasy, do której wpisywane są zwierzęta obu odmian barwnych oraz potomstwo powstające z ich krzyżowania między sobą.
W różnych krajach utrzymywana jest rasa holsztyńsko-fryzyjska, która zwykle powstała na bazie krajowej populacji krów krzyżowanych z buhajami holsztyńsko-fryzyjskimi (krzyżowanie wypierające). Dlatego obie odmiany barwne są zróżnicowane. Jeden z ekspertów z zakresu hodowli bydła podaje, że zaobserwowano w Polsce coraz częstsze wybieranie nasienia buhajów czerwono-białych, którym inseminowane są krowy czarno-białe. Jedną z przyczyn może być chęć zmniejszenia kalibru krów, ale też i poprawienie składu mleka. Jednak ze względu na zróżnicowanie bydła, tylko krowy z niektórych krajów wytwarzają mleko o lepszym składzie. Zanim podejmie się decyzję o zakupie nasienia, należy sprawdzić w katalogu buhajów średnią procentową zawartość tłuszczu i białka w mleku ich córek.
Choć potencjał produkcyjny europejskiego bydła czerwono-białego jest nieco mniejszy w porównaniu z czarno-białym i jest ono mniejszego kalibru, ale za to jest mniej wymagające i lepiej umięśnione. Trzeba jednak znowu zwrócić uwagę na zróżnicowanie i sprawdzić w katalogu buhajów wycenę budowy i pokroju córek danego rozpłodnika, ich wysokość oraz dolew krwi holsztyńsko-fryzyjskiej. Ekspert zwraca uwagę, że można spotkać buhaje czerwono-białe z dolewem 75% i mniejszym. Dobrym rozwiązaniem jest publikacja zdjęć córek buhajów.
Red factor
Popularne stwierdzenie mówi, że cel w stadzie można osiągnąć dopiero po trzecim pokoleniu, co odzwierciedla powolne tempo zmian genetycznych u bydła, choć nie dotyczy wszystkich cech.
W przypadku rasy holsztyńsko-fryzyjskiej za dominujące umaszczenie czarno-białe jest odpowiedzialny gen C, a za recesywne czerwono-białe – gen c. Na rys. 2 podano możliwe kombinacje genotypów i fenotypów powstające przy krzyżowaniu obu odmian barwnych rasy holsztyńsko-fryzyjskiej.
Jak widać, krzyżowanie buhaja czarno-białego z czarno-białymi krowami może dać w większości potomstwo czarno-białe, w jednym przypadku będzie ono czerwono-białe, ale część zwierząt pokolenia F1 będzie nosicielem genu c. Litera F, którą określa się pokolenia oznacza syna i córkę (łac. filius – syn, filia – córka).

Jeżeli krowy lub jałówki o genotypie Cc lub cC będą krzyżowane z buhajem czerwono-białym (cc), można uzyskać genotypy i fenotypy czarno-białe i czerwono-białe.
W katalogach buhaje czarno-białe będące nosicielami genu czerwono–białego umaszczenia bywają oznaczane symbolem RC lub RF (ang. red factor). Jeżeli publikowane są zdjęcia córek takiego samca, wówczas prezentowana jest jedna czarno-biała i jedna czerwono-biała.
Potomstwo czerwono-białe będzie można uzyskać wyłącznie w przypadku buhaja będącego nosicielem recesywnego genu umaszczenia czerwono-białego (RF) oraz czarno-białej krowy z takim genem lub krowy czerwono-białej.
Córki buhajów czerwono-białych, niezależnie od ich umaszczenia, będą miały cechy swoich ojców, czyli wzrost, umięśnienie i skład mleka dla odmiany czerwono-białej. Natomiast potomstwo czerwono-białe po buhajach czarno-białych z RF będzie w typie ojca czarno-białego.
Gen VCR
Dominujący gen czerwonego ubarwienia (ang. Dominant Red Carrier) ujawnił się w 1980 roku w Stanach Zjednoczonych u krowy rasy holsztyńsko-fryzyjskiej odmiany czerwono-białej, która w swoim genotypie posiadała dwa allele warunkujące barwę czarno-białą. Gen ten powstał w drodze mutacji.
Krzyżowanie wsteczne
Ten rodzaj krzyżowania pozwala ustalić nie tylko genotyp zwierzęcia w kontekście przynależności do rasy, gdy barwa jest cechą rasową, ale też służy do testowania nosicielstwa genów letalnych (śmiertelnych) powodujących np. śmierć zarodków, płodów i noworodków oraz semiletalnych obniżających żywotność, ale niekoniecznie prowadzących do śmierci.
Geny letalne są zmutowane w stosunku do genów normalnego rozwoju. Obecność takiego allela w genotypie heterozygoty może nie uwidocznić się, ale pojawi się w przypadku homozygoty recesywnej, czyli wówczas, gdy ten gen występował zarówno u krowy, jak i u buhaja.
***
Obecnie hodowca bydła i producent mleka mają do dyspozycji wiele narzędzi pozwalających ocenić stado pod względem wydajności, zdrowotności i genetyki. I choć popularne porzekadło mówi, że „krowa pyskiem doi”, wskazując jednoznacznie na żywieniowy charakter cechy ilościowej, jaką jest wydajność, to poznanie podstawowych zasad genetyki pozwala na jeszcze lepsze zrozumienie tego, co dzieje się w stadzie.
dr hab. Piotr Dorszewski, KPODR


